
Résumé :
Comment mieux préciser les relations entre l’individuation biologique, la biosphère et Gaïa ? Dans l‘ouvrage écrit par Lovelock en 1979, Gaïa et la biosphère sont clairement confondus. La tâche de cet essai est de trouver un argument théorique pour distinguer la géosphère de la biosphère, et pour mieux comprendre aussi les formes de tissage qui existent entre la géosphère et les organismes vivants qui la composent. Tout n’est pas dans tout, et il est important de trouver un argument pour mieux dissocier et connecter ces entités. Le point central sur lequel tout cet argument repose est la distinction que nous proposons d’introduire entre individuation physique et individuation biologique.
Mais en même temps nous voudrions trouver un argument compatible avec l’approche connexionniste et constructiviste proposée par Latour. Elle nous semble fournir une excellente perspective de recherche. Latour en effet refuse de tomber dans le Charybde du réductionnisme et dans le Sylla du holisme, pour traiter de Gaïa. Et cela exige de ne plus penser la biosphère et la géosphère simplement comme des sphères ! Il faut au contraire comprendre en quel sens on peut dire de la géosphère qu’elle est paradoxalement aussi une extension de la biosphère et non pas simplement ce dans quoi on peut déjà définir au préalable ses conditions de viabilité. La terre ne devient ainsi habitable pour les vivants que dans la géosphère. Dans la perspective que nous allons esquisser, la question de son habitabilité devient aussi une question de géophysique.
Introduction
Pour comprendre ce que dit Lovelock, insiste Latour, il faut commencer par sortir « d’une conception intenable de la totalité » (Latour, 2015, p 130). La véritable question qui se pose au sujet de Gaïa est : « comment suivre les connexions sans être holiste pour autant ? » (Latour, 2015, p 130). Et Latour d’ajouter ensuite, qu’il faut aussi « abandonner l’idée de partie » (Latour, 2015, p 131). Il invoque en effet l’hypothèse d’une pénétrabilité des entités proposée par Tarde et Whitehead. Gaïa n’est donc pas une totalité, au sens classique du terme. Latour ajoute que pour comprendre Gaïa, il ne faut pas se contenter de suivre les connexions, car Gaïa est aussi une « construction ». Pourtant, dans la quatrième conférence, Latour (2015) use d’un tout autre registre de vocabulaire. Il rappelle l’équivalence entre vie et autorégulation proposée par Lovelock pour définir Gaïa. Ce sont des mots que l’on trouve effectivement dans l’ouvrage de l’ingénieur anglais (Gaïa, a new look at life on earth ; 1979, p 11, p 49). En voici une citation rapide reprise et commentée par Latour dans son livre et dans un article écrit en réponse aux objections de Tyrell :
« I describe Gaia as a control system for the Earth – a self-regulating system something like the familiar thermostat of a domestic iron or oven » (1979, p 11).
Or s’il y a en effet un dispositif de contrôle, et même si comme pour tout dispositif de ce genre, sa logique est circulaire et non pas causale, n’est-ce pas forcément à un certain niveau qu’on peut remarquer sa présence ? Si on suit l’ingénieur anglais, tel semble bien être le cas. La terre a gardé une température constante, un taux d’oxygène et un taux de salinité des océans remarquablement stables que l’on ne peut pas expliquer par des considérations simplement physico-chimiques. Il semble que la biosphère se comporte ici comme une sorte de dispositif autorégulateur. Mais si on voit ainsi les raisons qui mettent en danger une approche réductionniste, on voit moins bien en quoi une approche holiste serait remise en cause. On ne voit pas non plus où et comment les termes de « connexion » et de « construction » pourraient trouver leur usage. Certes Lovelock parle d’un trial and error mechanism, mais il s’agit là d’un mécanisme pour expliquer la mise en place progressive d’un tel dispositif, qui, une fois actionné, garderait la même structure générale d’organisation.
Derrière ce premier problème, il y en a un second. Comment concevoir Gaïa, non pas comme un simple être vivant, mais comme une extension de l’organisation biologique ? Conscient des difficultés rencontrées dans sa première édition, Lovelock nous invite lui-même à suivre cette démarche :
« I failed to make clear that it was not the biosphere alone that did the regulating, but the whole thing, life, the air, the oceans and the rocks » (Préface de 2000).
Comment mieux préciser alors les relations entre l’organisation biologique, la biosphère et Gaïa ? Dans l‘ouvrage de 1979, Gaïa et la biosphère sont clairement confondus. Notre tâche est de trouver un argument théorique pour distinguer et articuler la géosphère de la biosphère, et pour mieux comprendre aussi les formes de tissage qui existent entre la biosphère et les organismes vivants qui la composent.
1- Pour une théorie de l’individuation biologique comme préalable.
La première étape de notre argumentation consiste à revenir sur l’usage que nous faisons du concept d’individuation biologique et des raisons pour lesquelles nous le préférons au concept d’organisation. Le terme d’organisation ou encore d’auto-organisation suggère simplement qu’un système est davantage en tant que tout que la somme de ses éléments. Mais le terme d’individuation utilisé en référence à son usage chez Gilbert Simondon (1964), désigne le fait qu’un système constitue un tout qui est en même temps élément de lui-même. Pour Simondon, en effet l’individu est paradoxalement le résultat de l’individuation. Il ne constitue donc pas une totalité close sur elle-même et il n’est pas non plus pourvu de parties au sens traditionnel. Or il y a de vrais points communs entre notre vocabulaire et celui utilisé par Latour (2012). Ce n’est sans doute pas un hasard puisque Latour est aussi un lecteur de Simondon, même s’il cite plus volontiers Tarde. Nous pensons donc que notre contribution peut aider à améliorer et expliciter la perspective qu’il propose.
* Tout d’abord, et comme Simondon lui-même, nous pensons qu’il existe déjà des systèmes physiques complexes dont on peut dire qu’ils sont éléments d’eux-mêmes[1]. Comment la localité d’un système se voit dans une transition de phase de premier ordre (par exemple la cristallisation) ? Nommons G la fonction qui régit les relations entre un système S et son environnement simplifié. G va par exemple avoir deux visages en fonction du fait que nous mettons en avant les variations de pression à température constante, ou les variations de température à pression constante. Mais dans tous les cas, et comme le dit Gibbs, G n’existe précisément, que dans la mesure où l’entropie de l’univers ne se mesure pas. G est simplement ce qui pilote la relation entre entropie et enthalpie dans une réaction chimique, ou encore une relation physique où il y a un échange avec le milieu extérieur. Qu’est-ce qu’un environnement simplifié ? Ce sont ce que les physiciens nomment des conditions limites, qu’il ne faut pas confondre avec les conditions initiales. Un système fermé a telle ou telle condition initiale. Il n’y a de conditions limites que pour les systèmes ouverts. Comment se voit alors la localité du système ? Elle se voit par le fait que pour certaines valeurs critiques de pression et de température, par exemple et s’il s’agit de transitions du premier ordre, les dérivées ∂G/∂P et ∂G/∂T de la fonction G ont une valeur infinie. Au voisinage du point critique, elles ne sont plus mesurables. Il y a deux dérivées, selon les deux visages de G :
∂G/∂P= V à T cte ; ∂G/∂T = S à P cte
La première est associée au volume, la seconde à l’entropie. Il en est de même si nous prenons un système ferromagnétique (transition de phase du second ordre). Les dérivées seconde de sa fonction G deviennent infinies. Qu’est-ce que cela signifie ? Ce que le physicien va analyser, c’est une difformité du système. Il ne se comporte pas dans son environnement, comme il se comporterait sans son environnement. Les équations de flux et l’action de son environnement sur lui ne sont pas uniformes. Elles sont difformes. Pour certaines valeurs des variables qui définissent cette action, une singularité, une non linéarité, ou encore une brisure de symétrie surgit. Nous pensons que cette action de l’environnement ne peut pas être décrite d’une manière mécanique. C’est une contrainte topologique. En d’autres termes, elle tient à la forme ouverte du système, et non pas à une cause, au sens traditionnel du terme.
Expliquons-nous davantage. On pourrait penser que G ne dit rien du système S de départ. Elle dit quelque chose de la relation entre S et son environnement. Nommons cela S* = S + E. Mais finalement ne décrit-on pas S* de manière complète ? Oui et non ! Pour décrire S*, il faut comprendre que S* est une extension de S. Et S, si on fait abstraction de son environnement, ne se comporte pas du tout comme S* ! Les difformités que les physiciens vont analyser, n’apparaissent en effet que dans S*. Elles n’apparaissent en aucun cas dans un système fermé, régi seulement par l’énergie libre de Helmholtz, et non par l’énergie de Gibbs notée G. Il y a donc une différence radicale entre S et S* que l’on ne peut pas comprendre, si on en reste à S. Cette différence n’est autre que la localité de S. Cette manière de concevoir la localité de S ressemble à ce que Latour appelle une « connexion » (Latour et alli, 2012). Nous ne pouvons jamais en effet traiter S comme un individu qui interagirait avec un autre individu. Il faudrait plutôt dire qu’il est interaction avant d’être individu. Ce que nous ajoutons simplement ici, c’est que cette notion n’est pas propre aux systèmes sociaux. Elle nous semble déjà avoir une pertinence pour les systèmes physiques complexes. Pour mesurer la localité de S, il faut toujours placer S dans quelque chose d’autre. Et nous allons donner un nom à cet opérateur de localité, ou encore de connectivité. Nous allons le nommer R1. Voilà pourquoi évidemment, tout comme il ne peut y avoir d’entropie de l’univers, il ne peut y avoir d’énergie libre de l’univers ! Telle est la leçon que nous tirons de la formule de Gibbs. C’est un principe fondamental de covariance (Van Fraassen, 1989) qui est ainsi annulé[2]. Le système S ne se comporte par comme l’univers, au sens minimal où il est justement impossible d’attribuer à l’univers les mêmes propriétés que lui. S a en effet pour propriété de connectivité R1 d’être à la fois un tout, une entité, et en même temps une propriété, ou un attribut qui apparaît dans la liste spécifiant une autre entité S* dont S dépend.
Il n’est guère étonnant ensuite que ce que nous nommons « R2 » puisse être associé à R1. Ce n’est du moins pas étonnant, du point de vue de l’ontologie du système dans lequel nous sommes ainsi placés. Dans certaines conditions et pour certaines valeurs de ses paramètres de contrôle, pression, température, susceptibilité, selon que la transition soit elle-même d’ordre un ou d’ordre deux, une propriété globale surgit. Ce que dit R2, ce n’est pas que le tout est plus que la somme de ses parties. Cela c’est justement la conception antique et insuffisante de la totalité, dont Lovelock reste partiellement prisonnier. Ce que dit R2, c’est que le tout est en même temps le résultat de ses opérations. Il est ainsi au sens strict et précis élément de lui-même. Il a un « auto », mais cet « auto » est pourtant, et très paradoxalement en même temps le produit de ses opérations. Il n’est donc pas individu. Il est individué. Dit en d’autres termes, on peut bien dire de R2 qu’il exprime le fait que le système a une dimension constructive.
En termes de physicien (R1 R2) signifient, que même s’il n’y a pas de solution analytique directe aux équations différentielles qui décrivent initialement les relations entre un système S et son milieu, en se plaçant dans un espace approprié, il est possible d’écrire une équation récursive de point fixe qui va nous donner la valeur de l’exposant critique permettant de mesurer exactement quand et comment une propriété globale émerge au sein de ce système S et qui le transforme en S*. Et cet exposant critique est toujours valable pour une classe de systèmes locaux et non pas pour un seul. Ce sont les techniques dites de renormalisation qui rendent cela possible. Mais cet espace approprié dont nous parlons n’est pas un espace d’états traditionnel. C’est un espace de modèles (Lesne, 2008). Il ne décrit pas l’univers comme un grand système dont on pourrait dégager la structure, à la manière de Laplace. Il décrit une propriété que nous observons à travers des modèles abstraits qui satisfont les données expérimentales que nous pouvons recueillir. Nous ne pouvons le dégager qu’en renonçant d’emblée à prétendre parler de l’univers, en tant que tel. Autrement dit, pour écrire les équations des groupes de renormalisation, il faut forcément intégrer à la fois la dimension de connexion et de constructivité des systèmes locaux que le physicien analyse.
** Mais ensuite, et comme beaucoup d’autres aujourd’hui (Bailly/Longo 2008, Soto/Sonnenschein 1999, Mossio/Moreno 2010, Montévil/Mossio 2015, Montévil/Pocheville/Longo 2012) nous pensons qu’il y a une différence d’ordre entre l’individuation physique et l’individuation biologique. Une telle différence d’ordre n’est pas au centre de la réflexion de Latour qui n’en évoque pas la possibilité. Elle nous semble pourtant tout à fait centrale pour éclairer le débat sur Gaïa et la biosphère. Elle va nous permettre en effet de présenter un argument pour éclairer les relations entre les organismes vivants et la biosphère, argument qui n’est pas présent dans le travail de Latour. Si l’on utilisait un vocabulaire bergsonien, il faudrait dire que cette différence d’ordre est une différence de nature et non pas une différence de degrés. Cette idée était tout simplement déjà celle de Gould. C’est dans « Full House » que Gould évoque ce qu’il nomme « the left wall of complexity ». Il y a un « mur de gauche » de l’individuation biologique, un peu comme au base-ball. Mais en quoi consiste cette différence de nature ? Nous proposons d’imaginer, à la manière de Simondon (1964), un système dont l’individuation est indéfiniment retardée, dans la mesure où elle est redoublée. Pour mettre en scène ce redoublement, nous proposons d’écrire l’équation récursive suivante, à travers laquelle les deux opérateurs R1 et R2 sont soumis à une condition R d’auto-application. Il s’agit évidemment d’une condition théorique. Ce n’est pas une condition empirique :
(1) R N = (R1 R2) N
Ce que nous imaginons est pourtant en adéquation avec les résultats des sciences expérimentales actuelles qui portent sur les origines de la vie. Elles montrent notamment en effet qu’il n’y a pas une, mais des origines. On ne peut plus dire par exemple que la vie trouve son origine dans la présence des acides aminés, ou qu’elle trouve son origine dans des gènes minéraux qui auraient préexisté aux gènes biologiques. Nous pouvons imaginer ainsi que la première contrainte émergente majeure soit de type structure dissipative, la seconde de type boucles de régulation catalytique positive et négative, la troisième la membrane, la quatrième le stockage de mémoire par certaines molécules, etc… A la limite N nous irions ainsi vers la contrainte de redoublement R qui n’est plus une simple contrainte du premier ordre. Et sous l’influence de cette contrainte, le système S, au terme des itérations que nous venons de décrire est transformé en B.
Que signifie le B que nous obtenons ? Cela signifie que le système en question n’est plus simplement élément de lui-même, comme un simple système physique. Il a une individuation redoublée. Sa structure devient duale ou bipolaire. En même temps qu’il est du monde physique, il est d’un monde d’individuation qui lui est propre. Il a une nouvelle carte d’identité. Nous allons supposer que ce monde tend vers lui-même à travers deux points fixes qui sont la localité internalisée du système B, et l’agentivité ou normativité internalisée du système B, de sorte qu’il y ait bien redoublement d’individuation :
(2) R = F1F2 R
Nous voyons que c’est du redoublement d’individuation R que nous supposons qu’il tend vers deux points fixes (F1F2) à travers lesquels sa structure d’individuation redoublée est le résultat de ses opérations. L’individu biologique est le résultat toujours provisoire de ce redoublement. Il n’en est en rien la cause. Telle est la supposition que nous formulons. Quelles en sont les conséquences ? Nous voyons aisément deux paramètres d’ordre, ou si vous préférez, deux propriétés émergentes apparaître en relation avec cette nouvelle équation de point fixe. Le premier n’est pas décrit par les analyses de Bailly et Longo. Il est par contre clairement formulé par Simondon.
Selon F1 le théâtre de normes qui caractérise un organisme vivant est toujours est constamment élément de lui-même, au sens où il ne forme un ensemble de contraintes que dans la mesure, où il dépend en même temps de contraintes qui ne sont pas les siennes[3]. Dit en d’autres termes, il n’est identique à lui, que dans la mesure où il diffère de lui, dans la mesure où il est spontanément, pour des raisons à présent internes (espace de contraintes) et non pas simplement externes à la fois en dedans et en dehors de lui-même, élément d’une niche biologique à travers laquelle il est co-construit (Lewontin, 2001). Il n’a d’identité que dans et à travers cette co-construction. Son identité est celle d’un nomade, et non d’un sédentaire. Nous voyons bien ce que cela signifie, puisque cela joue ici au niveau des contraintes. Cela ne signifie pas simplement que le système est local, comme un système physique. Cela signifie plutôt que sa localité est internalisée. Elle n’est plus de l’ordre du fait. Elle est transformée en norme. Sa localité vaut de droit, et le système commence ainsi spontanément à se donner son environnement et non pas simplement à le subir. Evidemment, une telle direction de pensée est exactement contraire aux postulats traditionnels du néodarwinisme. Si notre hypothèse est juste, plus un système biologique est individué, et plus il est co-construit. Cela se voit à un double niveau. Tout d’abord un système biologique n’est pas robuste au sens traditionnel du terme. Il est robuste en tant qu’il ne boucle sur lui-même comme ensemble de contraintes, que dans la mesure où il dépend en même temps de contraintes qui ne sont pas les siennes. Nous en avons une illustration parfaite dans le schéma proposé par Montévil/Mossio (2015).
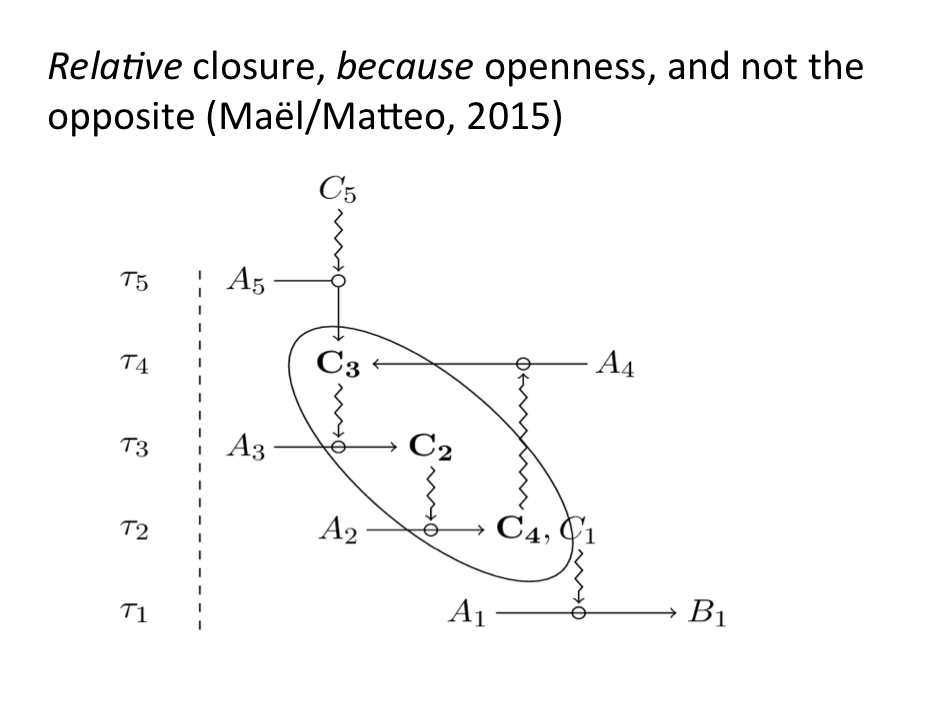
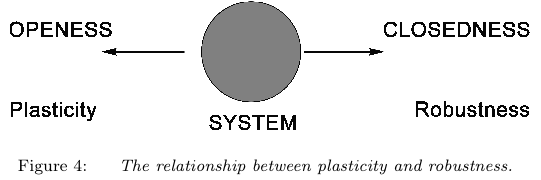
Ensuite, dans la perspective qui est la nôtre, il ne saurait y avoir de robustesse, sans qu’il y ait en même temps vulnérabilité et plasticité, précisément de par le fait que la localité d’un organisme est internalisée, cette localisation ouvre sur un potentiel de destruction et sur un potentiel de transformation.
Nous pensons que le cancer, le vieillissement ou la mort, sont des exemples de potentiels de destruction de ce genre. Il est évident dans cette perspective que le cancer est d’abord et avant tout une pathologie de l’individuation (Soto/Sonnenschein 1999) et non le résultat de la mutation d’un gène. Et, s’il y a une nécessité à ce qu’un organisme vieillisse ou meure, elle n’est pas intrinsèque au sens traditionnel du terme, elle est plutôt l’illustration du fait que sa localité est une norme, un réquisit. Il ne meurt pas. Il est un être pour la mort. Il ne tombe pas malade. Il est un être pour la maladie. Ce n’est pas simplement de l’humain qu’il faut dire cela, mais de tout être vivant.
Nous pensons aussi que l’internalisation de la localité est aussi ce qui fait de la plasticité une norme, de la plasticité, c’est à dire de la capacité de changer de normes en interaction avec son milieu. Cette capacité est pour nous le résultat de l’hétérogénéité propre à tout organisme vivant. Elle ne vient pas simplement des variations du milieu, comme si l’on s’en tient à la définition traditionnelle du terme en biologie du développement (West Eberard, 2003).
Par l’opérateur F2 nous supposons que l’agentivité ou la normativité d’un système biologique n’est à son tour pas un état qui lui est propre dans certaines circonstances. C’est une norme, une contrainte. Le vivant n’est pas simplement agentif (R2). Il est agentif de droit (F2). Il n’est pas simplement créateur. C’est une exigence pour lui d’être créateur. Nous pensons d’ailleurs que le concept de « Wille zur Macht » proposé par Nietzsche ressemble énormément à F2. La volonté de puissance n’est pas la volonté de quelqu’un. Ce n’est pas la volonté d’un individu, au sens traditionnel. Elle exprime plutôt un redoublement de l’individuation dont l’individu est l’effet, ou dans le langage de Nietzsche, un redoublement de la puissance. La puissance non plus comme simple état, mais comme tendance ou encore instinct.
Selon F2 et pour des raisons internes liées à l’espace/temps hypercritique dans lequel un organisme se constitue comme tel, sa singularité est générique, elle n’est plus spécifique. Cela signifie qu’un tel système ne connaît pas une transition critique dans certaines conditions et pour certaines valeurs de ses paramètres de contrôle. Nous supposons au contraire en accord avec les hypothèses de Longo et Montévil (2015) que sa criticité est internalisée. Il est donc de droit l’agent de ses transformations. Dit dans un langage plus philosophique, sa singularité est en même temps ce qu’il fait de ce qu’elle est. Sa singularité n’est pas un état, mais une construction dont il est en même temps ainsi rendu agent, il n’en est plus simplement l’objet. Il n’a rien de tout fait. Il se fait.
La stabilité d’un individu biologique existe. Il est élément de lui-même, il a donc un lui-même normatif. Il a une carte d’identité. Il a une anatomie et une physiologie. Il a une répartition du travail fonctionnel et physiologique. Les reins ne font pas circuler le sang. Je ne nie donc pas l’autonomie de l’individu théorisée par (Moreno/Mossio, 2010; Montévil/Mossio, 2015). Ils ont raison de la souligner. Mais ce « lui-même » est en quelque sorte toujours déposé par le processus de redoublement d’individuation que nous venons de décrire. Pour reprendre encore une formule de Nietzsche, il n’est qu’une petite raison toujours au service d’une raison plus grande. Il est un compromis entre ces deux archi-contraintes d’ordre topologique F1 et d’ordre chronologique F2. Sa stabilité est donc forcément relative, comme nous l’observons dans la réalité naturelle. Nous pouvons en faire des diagrammes. Mais ces diagrammes ne nous donnent pas directement les principes.
Nous pensons donc que l’individu est à chaque fois le résultat d’une forme d’interaction très complexe, mais dont la base fondamentale n’est autre que F1F2. L’un des éléments décisifs de cette complexité sur lequel nous allons insister dans ce papier est le découplage entre les trajectoires ontogéniques et phylogéniques, comme étant définitionnel d’un système biologique. Introduisons-le de manière frontale : il n’est pas possible de comprendre le dédoublement entre ontogénie et phylogénie, si on part de l’idée qu’un organisme est un ensemble de contraintes clos sur lui-même. Au contraire, si on admet que plus un organisme est individué et plus il est co-construit d’une part, et que d’autre part plus il est individué, et plus il est en mesure d’engendrer de nouvelles contraintes, le découplage de l’ontogénie à la phylogénie va forcément apparaître comme un puissant moteur d’individuation biologique ! Il introduit tout simplement une division qui garantit qu’un organisme ne saurait être reproduit à l’identique par sa descendance. Il est donc par sa phylogénie différent de soi, même si par son ontogénie il est au moins partiellement identique à lui-même. On voit ainsi que si un organisme n’est pas robuste sans être au moins partiellement plastique et vulnérable, cela pourrait bien être à cause de ce découplage entre phylogénie et ontogénie, à la frontière entre les contraintes développementales et les contraintes évolutionnistes. Ce découplage me semble une pièce maîtresse pour comprendre notamment en quoi un organisme peut être rendu autonome, au sens où il internalise son espace de contraintes (Mossio/Moreno 2010), tout en étant pourtant défini par le principe darwinien de descendance avec modification, qui est un principe non conservatif (Longo/Montévil, 2015).
2 - Gaïa et la biosphère
1- Peut-on dés lors concevoir Gaïa comme le résultat le plus global de ce processus de découplage entre ontogénie et phylogénie ? S’il y a une non identité à soi de Gaïa au sens technique que nous venons de donner, elle ne peut pas prendre la même forme que celle qu’elle aurait dans n’importe quel autre organisme biologique. On ne peut pas dire en effet de l’identité de Gaïa qu’elle est le résultat d’une co-construction, puisqu’il n’y a pas d’autres super-organismes avec lesquels Gaïa coopère. Gaïa semblerait alors se totaliser elle-même. Si une telle définition s’avérait juste, si on pouvait considérer la planète Terre comme un organisme autonome, au sens où il reproduirait sans cesse les conditions qui rendent possible son existence, pourquoi ne pas conclure d’elle qu’elle serait le seul véritable organisme vivant ?
Evidemment, dans une telle perspective, nous serions loin de la définition de l’individuation biologique que nous venons d’esquisser dans la partie précédente. Nous reviendrions plutôt à une définition de l’organisation biologique proche de celle proposée par Varela (1980) et ses disciples. Cela pourrait expliquer aussi que Gaïa puisse être sans contradiction un optimum et un thermostat. Dans l’univers de pensée néo-darwinien, cela n’a aucun sens. Mais dans la perspective de la clôture organisationnelle et de l’autopoièse, c’est tout-à-fait imaginable. Rappelons en effet que pour Varela et ses disciples, la structure d’un système biologique est en même temps le résultat de ses opérations, de telle sorte qu’elle est sans cesse reconduite à l’identique. Elle ne se transforme pas. C’est le concept de clôture organisationnelle. Si on comprend Gaïa de cette manière, qu’est-ce qui nous empêche de la caractériser comme un être vivant, si on admet par ailleurs que la clôture organisationnelle est une propriété fondamentale de la vie ?
Mais si au contraire, dans la perspective que nous défendons, il faut bien distinguer l’individuation biologique de la clôture organisationnelle, alors cela signifie que le découplage entre ontogénie et phylogénie est un élément fondamental pour définir un être vivant. Or, quelle forme pourrait bien avoir ce découplage pour Gaïa ? Gaïa n’a justement pas de descendance. Il n’y a pas de descendance avec modifications de la géosphère, pas même de la biosphère. C’est une confusion catégorielle qui nous a conduit à l’oublier. La descendance avec modifications ne vaut que pour des organismes vivants. Et Gaïa n’est donc pas un organisme vivant.
Par ailleurs, nous pouvons dire que Gaïa ne se reproduit pas, mais nous pouvons ajouter que Gaïa n’évolue pas non plus. C’est la biosphère qui évolue, ce n’est pas Gaïa. La biosphère, c’est-à-dire l’ensemble des organismes vivants. J’insiste, pour éviter toute confusion, dans la perspective que je défends ici, Gaïa et la biosphère ne sont donc évidemment pas identiques l’une à l’autre. Quant à la géosphère, c’est un ensemble de contraintes qui expliquent comment la température reste stable, comment le système d’autorégulation qui permet cette stabilité reste robuste. Autrement dit, c’est à nouveau la robustesse, qui caractérise la géosphère. Ce n’est ni la plasticité, ni l’évolvabilité ! Là encore, il semble qu’il y ait une confusion de catégories ; dont Lovelock est sans doute en partie responsable. Gaïa, c’est notre planète. Elle abrite la vie, mais la géosphère n’est pas la biosphère. Je définirai , en effet, la biosphère comme n’étant rien d’autre que la trajectoire phylogénique des êtres vivants, en tant que nous ne pouvons les comprendre qu’en refusant d’oublier le découplage fondamental entre phylogénie et ontogénie qui les définit. S’il y a quelque chose par excellence qui évolue, c’est bien la biosphère. Ce ne sont pas les organismes. Ces derniers s’adaptent, quand ils sont complexes, il se développent et montrent aussi leur vulnérabilité aux agents pathogènes. Mais ils n’évoluent pas ! Inversement commençons par nous demander à présent : est-il possible de concevoir la biosphère comme un super-organisme ? Même si elle ne se reproduit pas, même si elle n’a pas de descendance, n’est-ce pas la biosphère qui évolue ? Reprenons nos critères. Ne peut-on pas considérer la biosphère comme un théâtre de contraintes ouvert et normatif ? Précisons-mieux ce point. Peut-on vraiment dire que c’est la biosphère qui évolue, ou que ce sont les organismes vivants qui évoluent dans la biosphère ?
2- Il faut regarder de près le problème. Et pour le faire, nous allons provisoirement considérer la biosphère dans ses seules relations de composition avec les organismes vivants, et cela sans tenir compte pour le moment de la géosphère. La biosphère ne peut pas être un système biologique ouvert au même titre qu’un organisme vivant. Pourquoi ? La première réponse est que la biosphère forme un tout, mais elle n’est pas élément d’un autre tout biologique, au sens où elle ne fait pas partie d’une totalité biologique qui l’inclurait en elle. Mais la biosphère n’est-elle pas alors que globale ? Encore un coup comme éviter de faire de la biosphère une sphère ? Il y a là quelque chose de difficile à comprendre conceptuellement.
Posons le problème autrement. La biosphère n’est pas close, comme un thermostat ! Elle est bien ouverte, elle est même l’ouvert par excellence et le normatif par excellence, mais d’une manière différente d’un simple organisme. Elle est en effet plutôt la condition qui fait que tous les organismes présents dans la biosphère sont en même temps dans une niche, et qu’ils sont eux-mêmes des niches. Ils sont co-construits. Essayons d’analyser de plus près cette condition. En quel sens peut-on dire de la biosphère elle-même qu’elle est dans une niche, en même temps qu’elle est une niche ? En quel sens peut-on dire de la biosphère qu’elle n’est régulatrice, que parce qu’elle est régulée, autrement dit, qu’elle n’est un tout qu’en étant pourtant paradoxalement élément d’elle-même ? Parce que les éléments qui composent la biosphère sont en même temps eux-mêmes des totalités biologiques, des points de vue. Ce qui rend la biosphère structurellement locale vient des organismes vivants qui la composent. Cela ne vient pas du milieu biologique et physico-chimique qui l’environne. De la biosphère seule, on peut dire, qu’elle a complètement, entièrement son milieu en elle, au moins au sens biologique ! Voilà pourquoi elle continue en permanence d’évoluer. Elle est certainement l’entité biologique la plus radicalement détotalisée, puisqu’elle est détotalisée entièrement de l’intérieur ! Et c’est d’elle que nous pouvons dire en un certain sens qu’elle est le local et qu’elle est ce qui évolue par excellence, même si la biosphère ne se développe pas et ne prolifère pas, puisque toutes ses contraintes sont en elles. La biosphère est par excellence la condition d’internalisation de la localité dont tout organisme vivant dépend lui-même. C’est ce qui explique pourquoi il n’y a jamais de complète récapitulation de la phylogénie dans l’ontogénie. Dit dans le langage de Latour, les systèmes vivants ne sont pas simplement connectés, ils internalisent leur connectivité, et la condition d’internalisation n’est autre que la biosphère. C’est ce point à notre avis que notre analyse ajoute à sa brillante contribution.
3- Comment comprendre alors à présent les relations de composition entre la biosphère et la géosphère ? Une ambiguïté essentielle que l’on retrouve sans cesse à la fois chez Lovelock, mais aussi chez ses interprètes peu attentifs porte sur la question de savoir s’il faut considérer Gaïa comme un ensemble de conditions physico-chimiques rendant possible passivement la vie et son évolution, ou si au contraire il faut considérer Gaïa comme vivante ? C’est la logique binaire qui est à l’œuvre dans cette question que nous voudrions commencer par éviter. Il faut dire que l’ingénieur anglais a partiellement lui-même succombé à cette logique en acceptant d’abord l’analogie entre Gaïa et un organisme vivant.
Mais ce n’est pas le seul point troublant et approximatif dans son langage. Il affirme par exemple que partout où il y a de la vie, il y a un système qui prend de l’énergie libre à son milieu et qui rejette de la chaleur. Mais toutes les transitions de phase classiques du premier ou du second ordre, comme la cristallisation, le ferromagnétisme ou encore les structures dissipatives constituent des systèmes de ce genre ! Cette remarque ne nous permet donc en rien de distinguer l’individuation physique de l’individuation biologique. Elle ne nous permet pas de comprendre la différence entre un être vivant et un cristal. Revenons ensuite sur l’image du thermostat, telle qu’il l’utilise (Gaïa, a new look at life on earth ; 1979, p 11, p 49). C’est une image empruntée à la cybernétique. Un four moderne, affirme l’auteur, n’est pas un simple feu. C’est un dispositif doté de capteurs qui contrôle par régulation positive et négative la température du feu, de sorte qu’elle reste constante. On voit bien que la logique de ce dispositif n’est pas causale, elle est circulaire. C’est un dispositif autorégulateur. Pourtant encore une fois cette image est trompeuse. Dans un dispositif de ce genre, la circularité est engendrée à partir de la mise en place des capteurs par un ingénieur humain. Ce n’est en rien une propriété émergente du système ! La robustesse est obtenue artificiellement. Et nous avons vu qu’il existe une quantité de systèmes physiques robustes, qui ne sont pas de simples dispositifs artificiels, et au sein desquels au contraire une propriété globale émerge, de telle sorte que le système s’autorégule, car il est d’abord individué. Il est élément de lui-même en interaction avec son environnement. Tel est par exemple le cas pour les rouleaux de Bénard. Non seulement donc cette image est trompeuse, mais elle ne spécifie en rien Gaïa, en tant qu’en son sein, il y a intrication entre la géosphère et la biosphère. Pourtant nous allons séjourner un moment dans l’image et nous demander jusqu’à quel point elle peut néanmoins avoir une valeur heuristique. Et je pense en effet comme Latour qu’elle en a une.
Le point le plus frappant par lequel il faut commencer est sans doute la température de la terre. Qu’elle ait pu rester à peu près constante, ou en tout cas stable avec des fluctuations et donc robuste, voilà le point de départ du raisonnement de l’auteur. Je ne peux pas vérifier jusqu’à quel point c’est vrai. Mais cela ne semble pas démenti par les géophysiciens d’aujourd’hui. Certes on pourrait objecter, comme le fait Toby Tyrell (2013), qu’il s’agit peut-être d’un heureux concours de circonstances susceptible de changer à tout moment. Rien ne garantit a priori qu’elle soit structurellement stable. Mais qu’une planète dont la température est restée constante alors que son atmosphère était au départ chargée de méthane et à l’arrivée chargée d’oxygène, nous semble pourtant a posteriori constituer une marque troublante de robustesse ! Comme s’il y avait en effet à l’origine de cette stabilité une dynamique et une sorte d’attracteur. Bien entendu, aucun besoin d’évoquer l’intentionnalité et la cause finale pour cela. Ce point ne nous paraît même pas devoir susciter le moindre débat. Aurait-on l’idée de dire que puisque le climat est en partie modélisable grâce à l’attracteur de Lorenz, c’est que le climat est conscient ? C’est tout simplement absurde et stupide.
Ensuite et deuxièmement, le cœur du raisonnement de Lovelock n’est pas là. Il constate les différences énormes entre les atmosphères de Mars, de Vénus et de Gaïa. Et il remarque que celles des deux premières planètes sont plus proches de l’équilibre chimique que la dernière. Ce n’est pas le dioxyde de carbone qui domine sur notre planète. Il y a du nitrogène, du méthane mais aussi beaucoup d’oxygène qui devraient normalement réagir ensemble. Comment concevoir la stabilité de cette répartition loin de l’équilibre chimique ? En contribuant par exemple à enfouir le carbone dans les sols et à dégager de l’oxygène dans l’air, les organismes vivants ne participent-ils pas clairement de cette stabilité ? Ce point n’est d’ailleurs pas nié par Tyrell (2013). Au contraire. Mais c’est un premier point central pour Lovelock, car cela signifie que ce sont des paramètres biologiques qui semblent ainsi être à l’origine de cette stabilité loin de l’équilibre du système géophysique. Et cela lance d’ailleurs tout un programme de recherche. On peut notamment citer la mise en évidence du rôle du dimethylsulfure (DMS) produit par une variété d’algues et qui est largué, puis oxydé dans l’atmosphère en contribuant ainsi à la formation des nuages et à son refroidissement. On voit ici la subversion du raisonnement pyramidal traditionnel à l’œuvre dans la pensée anglo-saxonne ! La stabilité géophysique ne nous donne pas une structure de base à partir de laquelle il faudrait se demander, s’il y a ou non émergence de propriétés biologiques, de telle sorte que le réductionniste puisse répondre « non » et que le holiste puisse répondre « oui ». Le réductionniste et le holiste ont tort tous les deux, car la question n’est tout simplement pas bien posée !
Formulée de cette manière, elle repose sur un postulat commun : la vision pyramidale de la Nature héritée de Lloyd Morgan (1923). Dans le raisonnement de Lovelock, la géochimie est au contraire courbée par la biologie. Ce n’est pas la biologie qui repose sur une base géochimique. La stabilité physico-chimique illustrée ici par la constance de la température de la terre n’est rien d’autre qu’une construction. Elle résulte des interactions à l’œuvre entre les entités biologiques et leur support physico-chimique. C’est cela qui nous empêche de penser Gaïa comme un simple support. Mais en même temps rien dans cette approche ne vient invalider l’idée que Gaïa pourrait être un être vivant. Ni que Gaïa se résume à un système autorégulateur, comme en cybernétique. N’oublions pas que les contresens que Bruno Latour attribue aux détracteurs de Lovelock, sont d’abord des contresens qu’il a fait lui-même et sur lesquels il est ensuite revenu.
Allons plus loin, rejetons le modèle cybernétique. Acceptons que Gaïa ne puisse se comprendre simplement de cette manière. Rien dans les analyses que nous venons de mener ne nous conduit à rejeter la thèse selon laquelle Gaïa serait bien l’entité biologique par excellence définie par une forme de clôture organisationnelle, au sens de Varela et de ses disciples. Gaïa serait ainsi un système de contraintes qui s’auto-reproduit continuellement, à condition de bien comprendre la différence entre les flux géophysiques et les contraintes biophysiques, comme dans le modèle de Varela. Or c’est cela que nous contestons ! Nous avons vu qu’il faut caractériser un être vivant par l’individuation et non par l’organisation ! Cela signifie qu’il n’y a pas de vie sans découplage entre l’ontogénie et la phylogénie. Or ce découplage n’a évidemment aucun sens pour Gaïa et le raisonnement ne marche donc pas ! Comment sortir du dilemme. C’est Lovelock qui va nous aider. Rappelons qu’il écrit en effet :
« I failed to make clear that it was not the biosphere alone that did the regulating, but the whole thing, life, the air, the oceans and the rocks » (Préface de 2000).
Nous allons à présent proposer une lecture de cette phrase en phase avec celle qui est suggérée par Latour. Gaïa n’est ni une simple condition pour l’émergence de la vie, ni une manière de dire que la terre est vivante. Gaïa est une forme d’extension constructive de la biosphère. Voilà aussi pourquoi Gaïa et la géosphère ne sont pas synonymes. Telle est du moins la thèse que nous défendrons ici.
Les logiciens distinguent dans les systèmes formels deux formes d’extension. Imaginons un système formel A. Supposons maintenant que nous puissions lui donner tout un ensemble de modèles A* qui le satisfont. A*contient des modèles qui sont non isomorphes à A. Pourtant chaque modèle présent dans A* est compatible avec la structure axiomatique de A et avec l’ensemble des théorèmes que l’on peut déduire de cette structure. C’est une extension conservative. Le schème pyramidal s’applique. A est la base et A* l’ensemble des modèles que l’on ne peut réduire à cette base, mais qui en dépendent pourtant essentiellement. Mais il existe aussi des extensions constructives ! C’est notamment Gödel qui a démontré l’existence de telles extensions. Prenons un système PA. Il y a une extension constructive ZF de PA si on peut démontrer dans ZF une formule qui existait dans PA sans qu’on puisse la démontrer dans PA. Cela vient évidemment du fait que la structure de PA est ouverte et que l’on peut démontrer dans PA qu’il existe des formules, dont on ne peut décider à l’intérieur de PA, si elles sont démontrables ou non démontrables !
Gaïa ne nous semble être rien d’autre que cela. Elle émerge de la biosphère comprise comme un système évolutif, mais qui est pourtant en même temps, une fois constitué, en interaction avec l’air, l’océan et les rochers qui ne sont justement pas des êtres vivants. Autrement dit, Gaïa est ce qui nous force à admettre une mixité des entités, comme celle que Latour (2016) appelle de ses vœux. D’un premier point de vue, la nouvelle entité « biosphère » forme un tout découplé des éléments biologiques qui la composent et en relation de connexion avec eux, comme nous l’avons vu. Mais d’un autre point de vue, elle dépend à présent rétrospectivement des relations entre les êtres vivants qui la composent, et de l’air, de l’eau, du feu, des rochers et des montagnes de la terre. Tel est le postulat qui est au centre d’un raisonnement constructif, selon Latour. Prenons le temps de citer son travail :
“Step 1, choose an entity A to start with – a phenomenon like bacteria respiration, or crustal rock weathering; step 2, shift attention to its surroundings (precisely, as we shall see, what Tyrrell, intoxicated by the metaphor of ‘selfish genes’, forbids himself to do); step 3, detect in those surroundings what transformation the entity A has induced; step 4, detect in those surroundings what transformation they have on A; step 5, compound the reciprocal effects by a gross use of the notion of negative or positive feedback, not because you believe there is a machine and an engineer (more of this later), but just to make sure the two are ‘closely coupled’; step 6, a tricky step, now, choose this ersatz of a feedback loop as the new starting point; step 7, start again so that ‘entity plus surroundings’ are now replaced by loops interfering with other loops; step 8 (the most important one in my view), anxiously revise the description so as to make sure the loops upon loops are not added to one another as if they were one Whole above the entities you started with.” (Latour, 2016, p 12)
La terre, les océans, le feu, la mer et l’air réagissent sur les êtres vivants et sur la biosphère, en même temps que cette dernière à présent les contrôle. Il n’y a pas de simple hiérarchie avec une totalité totalisée, il y a Enchevêtrement, Mixité des entités, il n’y a pas simplement un nouveau tout qui émerge en plus de ses éléments. Il y a connectivité. L’émergence de la biosphère fait aussi que la géosphère recompose la biosphère en même temps qu’elle est réciproquement recomposée par elle. Car en effet la biosphère est élément d’elle-même dans la géosphère, en même temps qu’elle l’est par rapport à chaque organisme vivant, mais pas sur le même mode, car la géosphère n’est pas en vie. C’est ainsi que nous pouvons tisser et articuler, mais sans tout mélanger.
En un sens Gaïa est moins que la biosphère, car elle n’est qu’un ensemble de paramètres physicochimiques. Mais en un autre sens elle est plus ! Elle est plus car elle est la marque que la géosphère est non seulement courbée par la vie mais peut-être même aussi rendue ainsi habitable[4]. C’est cela la Terre. C’est ce mixte, c’est cette marque. C’est cette subversion de la vision pyramidale de la nature, avec au premier étage la physique et la chimie et au deuxième étage la biologie. Il n’y a pas de maison et il n’y a pas d’étage, car la nature n’est pas humaine ! Comme le montre si bien Montebello dans son dernier livre (2015) la nature n’est pas là pour nous, et Gaïa en est justement la preuve. C’est en ce sens que nous ne voyons pas d’objection à dire que Gaïa contraint la physique et la chimie de la terre à se plier aux contraintes propres à la biosphère, mais dans l’exacte mesure pourtant où la géosphère la contraint aussi en rendant la terre habitable. L’habitabilité de la terre, propriété pourtant normative, ne relèverait pas que de la biosphère. On pourrait l’envisager ainsi comme une sorte d’extension constructive.
Pourtant, contrairement à d’autres, nous ne pensons pas que cette mixité rende Gaïa réfractaire à tout forme d’explication scientifique. La constance de la température de la terre, le taux de salinité de la mer, le taux d’oxygène de l’atmosphère qui reste stable un milliard d’années, tous ces éléments associés à la présence d’un radiateur permanent autour duquel notre planète tourne forment un ensemble très troublant. C’est comme si nous avions là un très spécial système thermodynamique ouvert et loin de l’équilibre, avec des paramètres de contrôle (température, luminosité), et une série d’exposants hypercritiques garantissant probablement de manière stable la plasticité évolutive de la biosphère et en même temps pourtant construits par elle, émergeant d’elle en interaction avec la géosphère! C’est comme si nous avions là sous les yeux les mystérieux exposants qu’on pourrait rêver de rattacher un jour aux origines de la vie ! C’est comme si sous nos yeux se manifestait la limite abstraite à travers laquelle les concepts de mixité et d’enchevêtrement deviennent opératoires !
Conclusion
* Si on part de la méthode traditionnelle en considérant qu’il y a d’abord des objets ou des agents, puis qu’il faut observer les interactions entre eux, mais aussi les interactions entre eux et leur milieu, pour finalement dégager la structure du système qu’on analyse, tout ce que nous venons d’écrire est obscur, voire incompréhensible. On peut se disputer alors pour savoir par exemple si la géosphère a des propriétés structurales qui ne se réduisent pas à la somme des éléments qui la composent ou non. On tombe dans le Charybde du réductionnisme et le Silla du holisme. C’est l’approche traditionnelle. Elle n’apporte rien au problème que nous posons.
Mais dans l’approche que nous proposons, en un sens la biosphère trouve son extension dans la géosphère, parce qu’en un autre sens la géosphère peut aussi être considérée comme une extension de la biosphère. Nous n’avons pas simplement affaire à des agents ou à des entités, mais à des points de vue singuliers. Et l’une des caractéristiques de ces points de vues qui ressemblent à ce que Tarde et Latour nomment des monades, c’est que l’un d’entre eux peut tout aussi bien entrer dans la liste des propriétés et des attributs qui en caractérisent un autre. Ce qu’il nous faut chercher alors, n’est pas comment en ramener l’un à l’autre, mais les recoupements entre elles (overlapings), les classes d’universalité, qui permettent de les articuler l’un sur l’autre. Gaïa nous semble être un excellent candidat, à l’intermédiaire entre biosphère et géosphère.
**Ensuite, nous pensons qu’il n’est pas simplement possible de chercher les modes de connexion entre ces points de vue singuliers, mais aussi les conditions particulières qui font qu’une forme de mixité, comme Gaïa_ qui n’est donc ni simplement la biosphère, ni simplement la géosphère, mais leur connectivité_ est en mesure d’émerger. Pour les trouver, il nous semble que la méthode de recoupement proposée par Latour ne suffit pas. Elle est trop descriptive. Nous croyons que ces conditions existent et que les savants de demain vont bientôt les découvrir. Nous pensons qu’un élément clé pour le comprendre, se situe au niveau de l’individuation biologique qu’il ne faut pas confondre avec l’organisation, au sens de Varela. Et nous pensons que cet élément n’est autre que la condition R qui explique pourquoi l’individuation biologique n’est pas une individuation comme les autres et qui éclaire la relation entre les organismes vivants et la biosphère. C’est aussi un argument central pour comprendre en quoi Gaïa ne peut pas être considérée comme vivante et en quoi il existe encore une distinction importante entre biosphère et géosphère qu’il faut maintenir.
*** Enfin dans tout le raisonnement ici exposé, le lecteur attentif ne manquera pas de noter que nous n’avons eu recours qu’au principe de descendance avec modifications et au découplage entre ontogénie et phylogénie pour caractériser la biosphère. Nous n’avons eu besoin en rien de faire intervenir le principe de sélection naturelle… Ce principe ne nous semble pas plus clairement attesté en biologie que le principe d’utilité en économie, et cela malgré la quantité impressionnante de papiers qui lui sont consacrés. Mais peut-être sommes-nous dans l’erreur ! L’histoire le dira. Enfin, en ce qui concerne le second, il nous semble que le théorème d’impossibilité d’Arrow a déjà largement rendu son verdict, même si bien peu nombreux sont ceux qui en connaissent l’existence.
Nous n’avons pas eu besoin non plus de dire que le vivant se reproduit. La reproduction nous paraît être un lointain héritage aristotélicien qui n’éclaire pas, mais obscurcit au contraire le bon usage du principe de descendance avec modification au niveau phylogénique, et du principe de prolifération avec variations au niveau ontogénique (Soto et alli, 2016). Mais sans doute sommes-nous aussi dans l’erreur !
- [1] Dans un système physique complexe, la localité du système se voit. Pour reprendre la belle expression de Latour, il faut détruire partiellement l’image du « globe » pour le comprendre. Il existe plusieurs types de systèmes physiques complexes. Les analyses récentes de Van Frassen (2004) et Bitbol (2007) montrent amplement qu’un système quantique est déjà complexe en ce sens. Quand on affirme la complétude structurale d’un système quantique, on oublie toujours que c’est dans un cadre qui est celui des inégalités de Heisenberg. On va toujours pouvoir décrire les états quantiques d’un système dans une certaine langue, à condition toutefois que nous admettions qu’il y a plusieurs langues pour les décrire, et que nous ne pouvons pas toutes les parler à la fois. Dans une certaine langue, une observable est aussi la valeur propre d’un vecteur propre projeté un certain nombre de fois sur lui-même, parce qu’en même temps dans une autre langue, tel n’est pas le cas. Et cette impossibilité de parler en même temps ces langues est elle même inscrite dans le formalisme, par le jeu croisé des transformées de Fourier qui est le principe secret sur lequel repose les inégalités de Heisenberg (voir notamment Lévy-Leblond/Balibar, 1987, pp 236-137).
- [2] « The equation is covariant if it is either true for all the frame of reference or for none » (1989, p 281).
- [3] Dit en d’autres termes, c’est un ensemble qui est toujours en même temps élément d’un autre ensemble. Il est structurellement potentiel. Il n’est pas actuel.
- [4] Voir Dutreuil S (2014).
Lectures
Bailly F, Longo G (2008) Extended critical situations. Journal of Biological Systems 16(2): 309–336.
Bitbol M (2007) Ontology, Matter, Emergence, Phenom Cogn Sci, 6 pp 293–307.
Dutreuil S (2014) What good are abstract and what-if models? Lessons from the Gaïa hypothesis, History and Philosophy of the Life Sciences Volume 36, Issue 1, pp 16–41.
Gould SG (1996) Full House, New York, Harmony Books.
Kauffman, S. (2000). Investigations. Oxford University Press.
Latour B (2015) Face à Gaia, Paris, La découverte.
Latour B (2016) Why Gaia is not a God of Totality, Theory, Culture, Society, Sage publication.
Latour B, Jensen P, Venturini T, Grauwin S and Boullier D (2012) ‘The whole is always smaller than its parts’ – a digital test of Gabriel Tardes’ monads, The British Journal of Sociology 2012 Volume 63 Issue 4, pp 590-615.
Lesne A et alli (2008) Physique statistique. Chaos et approches multi-échelles, Paris, Belin.
Lévy-Leblond JM, Balibar F (1987) Quantique, rudiments, Paris, Dunod.
Lewontin R. (2001) Gene, Organism and Environment: a new introduction, in Cycles of Contingency, Oyama. S, Griffiths P. and Gray R.D. editors, MIT Press.
Lloyd Morgan (1923) Emergent Evolution, Henry Holt and Co.
Longo, G., Montévil, M. & Pocheville, A. (2012). From bottom-up approaches to levels of organization and extended critical transitions. Frontiers in Physiology, 3, 1-11.
Longo G, & Montévil, M. (2014). Perspectives on organisms, Biological time, Symmetries Singularities. Berlin Heidelberg, Springer.
Longo G, Montévil M, Sonnenschein C, Soto AM In Search of Principles for a Theory of Organisms, in press, J. Biosciences, 2015
Lovelock J ( 1979) (2000) Gaia: A New Look at Life on Earth, Oxford University Press.
Miquel PA (2015) Sur le concept de nature, Paris, Hermann, Vision des sciences.
Miquel PA, Hwang SY (2016), From physical to biological individuation, Progress in Biophysics and Molecular Biology, Available online 16 July 2016, ISSN 0079-6107
Montebello P (2015) Métaphysiques cosmomorphes, Les presses du réel.
Montévil M, Mossio M (2015) Biological organisation as closure of constraints Journal of theoretical biology, 372: 179-192.
Mossio M, Moreno, A (2010) Organisational closure in biological organisms, History and philosophy of life sciences 32 : 269-288.
Simondon G (1964) L’individu et sa genèse physico-biologique, PUF, Paris.
Soto A, Sonnenschein C (2004) Emergentism as a default: Cancer as a problem of tissue organization; J. Biosci. 30: 103–118.
Soto A, Longo G, Miquel PA, Montevil M, Mossio M, Perret N, Pocheville A, Sonnenschein C (2016) Toward a theory of organisms: Three founding principles in search of a useful integration, Progress in Biophysics and Molecular Biology, http: //dx.doi.org/10.1016/j.pbiomolbio.2016.07.006.
Tyrrell T (2013) On Gaia: A Critical Investigation of the Relationship between Life and Earth. Princeton: Princeton University Press.
Van Fraassen B (2004) Science as representation: flouting the criteria Philosophy of Science 71:794–804.
Varela F (1980) Principles of Biological Autonomy, North Holland/Elsevier, New York.
Varela F( 1981) Autonomy and Autopoiesis, Self-organizing systems - autopoiesis.com.
West-Eberhard M.J. (2003). Developmental Plasticity and Evolution. Oxford University Press.